
刘小坤团队在植物学权威期刊《Plant Biotechnology Journal》发文综述植物-病原菌拮抗操控糖转运蛋白竞争糖类的研究进展

在生态系统中,植物作为自养生物,借助光合作用将阳光转化为能量,合成糖分以供自身使用。然而,植物病原菌作为异养生物,必须从寄主植物中汲取糖分才能完成其生命周期,这便引发了植物与病原菌之间围绕糖分资源的竞争。由于植物体内糖分资源有限,这种竞争变得尤为激烈。病原菌需要与植物争夺有限的糖分以维持其生存和繁殖,而植物则采取措施限制病原菌获取糖分,从而阻止其进一步侵害。因此,植物与病原体之间如何巧妙地调节糖分供应,成为决定抗病性或易感性的关键因素。
近日,庐山植物园的刘小坤研究团队与英国Tony Miller教授团队合作,在国际权威期刊《Plant Biotechnology Journal》上发表了一篇题为“The role of sugar transporters in the battle for carbon between plants and pathogens”的综述文章。该文系统回顾了糖转运蛋白在植物与病原菌相互作用中的研究进展,探讨了双方如何通过调控糖转运蛋白,以争夺宝贵的糖分资源。

当病原菌侵袭植物时,糖分通过两条主要途径从远处组织输送到感染区域:一是质外体途径,糖分由转运蛋白从远处运至质外体,随后通过扩散到达感染点;二是共质体途径,糖分通过胞间连丝和细胞膜构成的通道直达感染部位。病原菌依据其种类、生活方式以及感染阶段的不同,采用多样化的机制吸收糖分。
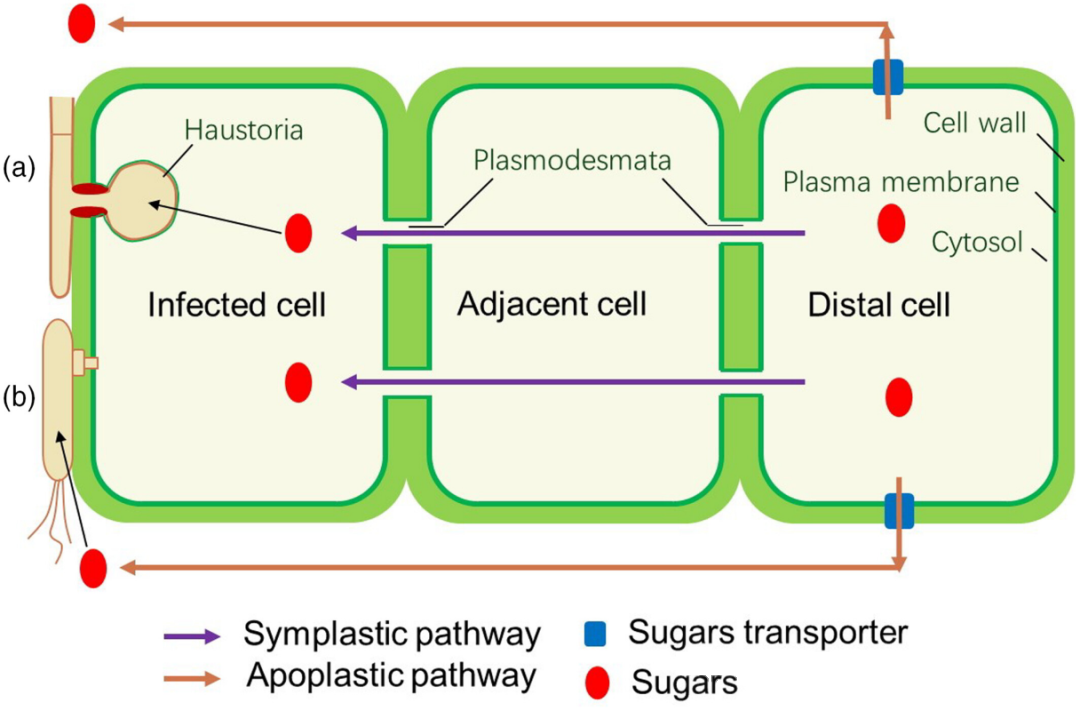
图1. 植物与病原菌互作时,植物体内糖类移动的两种途径
植物体内存在三类主要的糖转运蛋白:SWEETs、SUTs和STPs。它们分布于特定的亚细胞膜上,具有特定的糖底物和运输功能。植物通过这些糖转运蛋白与转化酶invertase协同作用,调控糖分的运输、分配和供给,响应内外部信号变化。例如,蔗糖可被SWEET蛋白运至质外体,再由SUT蛋白回收入细胞;或者在细胞外被糖转化酶分解为单糖,之后由STP蛋白重新摄入细胞。此外,液泡膜也参与糖分的运输与分配过程,如液泡内的蔗糖可被糖转化酶分解为单糖,并由ERD6排出至液泡外的细胞质中。
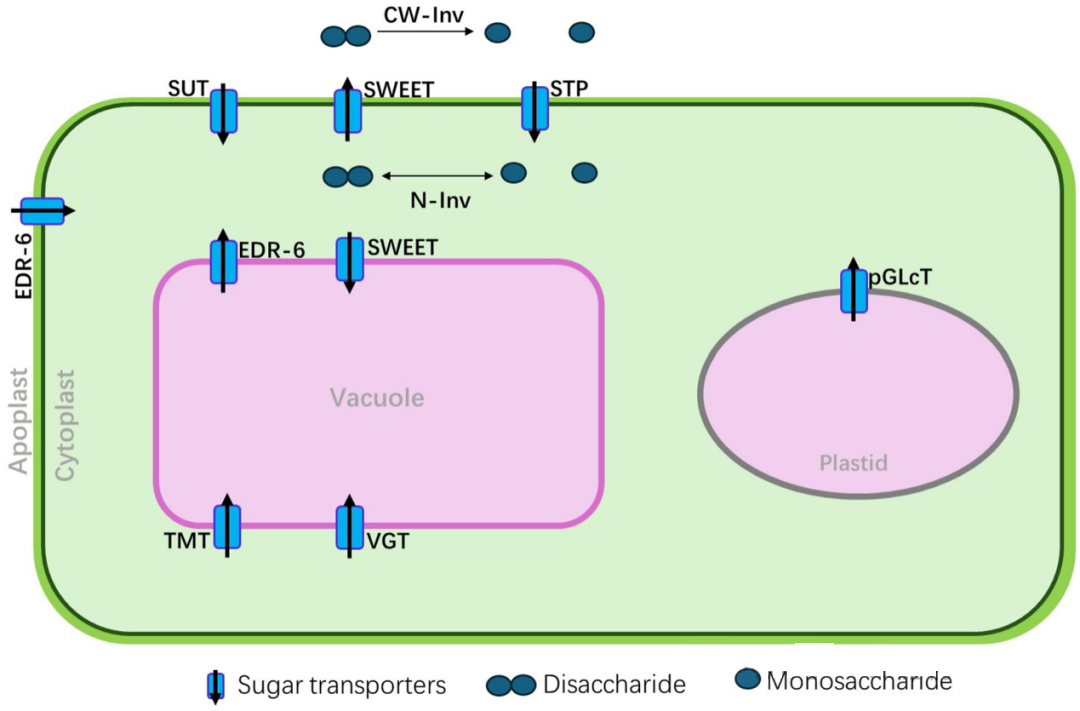
图2. 植物体内的各种糖转运蛋白和功能
进化研究表明,SWEET和STP蛋白不仅存在于真核生物中,还出现在原核生物中,而SUT蛋白则仅限于植物和真菌。某些植物中,糖转运蛋白经历了快速进化,表明其功能可能更为多样化。在植物与病原菌的互动中,植物的转运蛋白受到双方的拮抗性调控,影响抗病性或易感性。例如,对于活体营养型真菌而言,病原菌感染促使STP蛋白将单糖吸入细胞,供给吸器使用,促进感染进程;然而,当Lr67res蛋白(STP同源蛋白)感染时,该蛋白功能丧失,无法将单糖摄入细胞,从而产生抗病效果。对于细菌病原体,其通过分泌TAL效应蛋白上调SWEET蛋白的表达,增加细胞外糖浓度,有利于细菌生长;而当植物表面受体检测到flg22信号时,则激发下游BAK1磷酸化STP13,增强细胞外糖的吸收,提高抗性。死体营养型真菌Rhizoctonia solani可通过诱导OsSWEET11表达,导致植物易感;反之,过表达STP13则能增强对死体营养型真菌B. cinera的抵抗力。
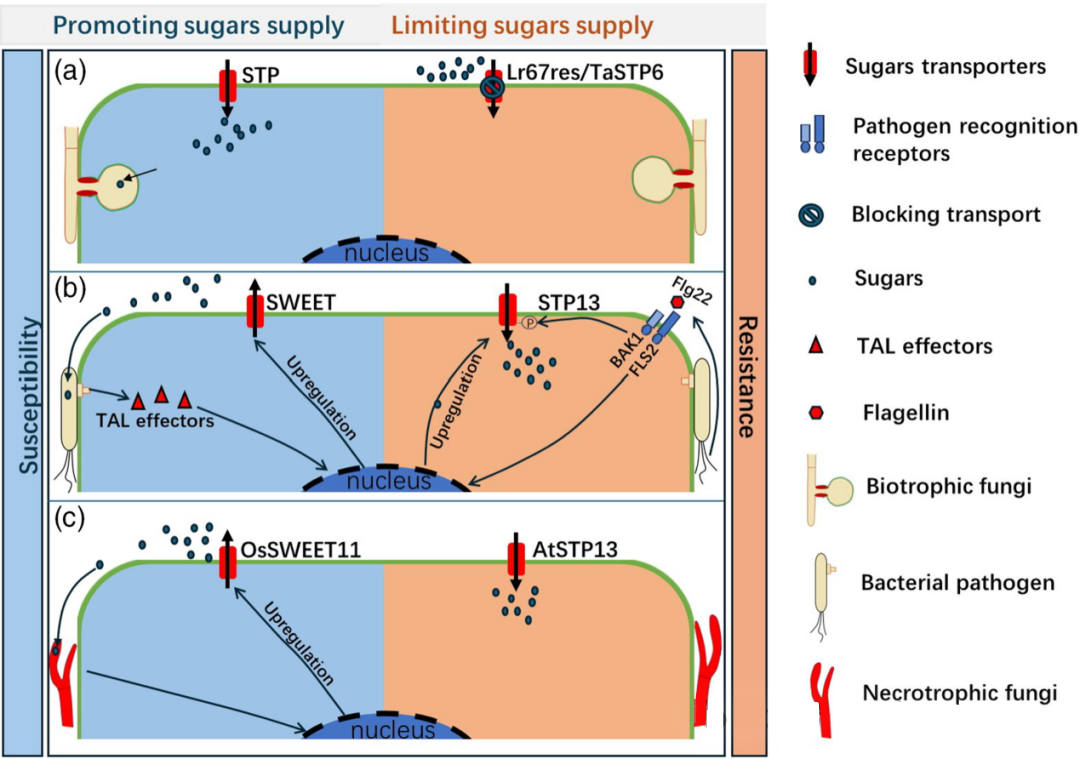
图3. 植物-病原菌互作中的糖转运蛋白的角色
除植物糖转运蛋白在植物-病原菌互动中的核心作用外,病原菌亦演化出独特策略,利用糖转运蛋白和糖转化酶与植物竞争糖源:提高与底物的亲和力以竞争糖分;通过糖转化酶干扰植物糖分配;利用罕见糖作为底物避免直接竞争。
当前,基于对糖转运蛋白在植物病害互作中角色的理解,抗性育种研究已取得一定进展,主要策略包括:通过持续遗传修饰糖转运蛋白限制病菌糖源,产生抗病性;通过特定时空依赖的遗传修饰糖转运蛋白,实现抗病性;筛选自然变异的转运蛋白,限制病菌糖源,达到抗病效果。每种策略都有其利弊,关键在于平衡植物抗病性和生长需求。
植物为抵御病原菌,发展出多元化的防御机制:一是识别病原体并激活免疫反应清除之;二是切断病原菌的营养来源,使其因饥饿而无法生存,进而形成抗性。结合这两种策略,可能为我们提供新机遇,开发出更加持久且高效的抗病植物品种。
庐山植物园的刘小坤研究员为通讯作者,约翰英娜斯中心的Chen Yi博士为第一作者,约翰英娜斯中心Tony Miller教授,庐山植物园的裘博文,南昌大学的黄瑶,厦门海洋三所的张凯博士,厦门绿化中心的Gaili Fan博士参与了本项研究。该研究得到了江西省自然科学基金,九江市科学自然基金和江西省千人计划创新领军人才长期项目的资助。


